




© Mario Izquierdo
Los organismos procariontes poseen unas características que los diferencian de los eucariontes:
- Poseen un tamaño entre 1 μm y 5 μm, con flagelos de estructura fibrilar o movimiento deslizante, mientras que los eucariontes miden más de 5 μm.
- La pared de los procariontes posee varios polímeros, casi siempre de un compuesto llamado 'peptidoglicano', que no se encuentra en las células eucariontes.
- A diferencia de los eucariontes, no poseen vacuolas, y si las tienen, suelen estar llenas de gas.
- Los procariontes no poseen membrana nuclear, y solo tienen un cromosoma circular, que se encuentra unido a la membrana celular, o bien incluido en invaginaciones de la membrana celular, denominados 'mesosomas'.
- La replicación del ADN y su segregación son bidireccionales, con un sólo origen de replicación, siendo la segregación amitótica.
- En la síntesis de proteinas la traducción y transcripción se realizan al unísono.
- Los ribosomas son más pequeños (70 s de velocidad de sedimentación), y son inhibidos por antibióticos específicos de los procariontes.
- En estos organismos la producción de energía se realiza por fermentación, respiración o fotosíntesis. La cadena respiratoria se asocia a la membrana plasmática, y la fotosíntesis se realiza en estructuras que son invaginaciones de la membrana.
- La reproducción es asexual, por escición. Raramente existe conjugación no asociada con la reproducción y origina diploides parciales.
- Los procariontes se suelen dividir en dos filos:
| Schizomiceta, que incluye a las bacterias. | |||
| PROCARIONTES (MONERA) | Cyanophyta, que incluye a las cianofíceas | ||
| o algas verdeazuladas | |||
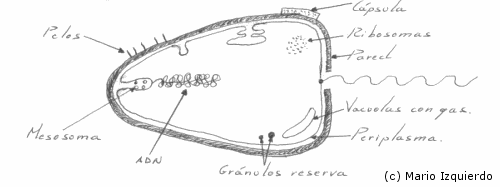
Muchos microorganismos procariontes presentan una capa externa mucilaginosa, denominada 'cápsula', que rodea completamente al organismo y cubre la pared celular. El tamaño de la cápsula y la cantidad de material producido depende de las condiciones del medio, o el cultivo en el que se mantienen. Algunas especies carecen de cápsula y sólo se forma un mucílago independiente de la célula.
La cápsula y el mucílago se componen, en general, de polisacáridos, conocidos como extracelulares, para diferenciarlos de los compuestos que forman parte de la pared celular.
En estos compuestos se han diferenciado muchos monosacáridos (Azúcares neutros, aminoazúcares y ácido murámico). Una cápsula puede proteger al organismo contra la secación, la infección por un bacteriófago o fagocitosis. En las bacterias patógenas, la formación de cápsula va asociada con el grado de virulencia, porque la cápsula protege del ataque de anticuerpos y macrófagos.
En la mayor parte de las bacterias, la forma de la célula se debe a la presencia de una pared celular rígida, cuya función es proteger al protoplasma que se compondría de la membrana celular y su contenido, esta pared celular protege al protoplasma de la lisis osmótica. Normalmente los organismos viven en un medio hipotónico con respecto al citoplasma. La rigidez de la pared celular se debe sobre todo a un sólo polisacárido, aunque existen otros polisacáridos característicos de cada grupo de procariontes, e incluso de las estripes que pertenecen a una misma especie. Los polímeros que forman la pared celular se organizan en estructuras complejas de varias capas.
La pared posee además otras funciones, como por ejemplo, los polímeros muy cargados pueden facilitar un mecanismo de intercambio de electrones, que colabora en la captura de sustancias vivientes.
La pared es un punto celular muy activo, que impide la entrada de algunas moléculas, y retiene proteinas en el periplasma (zona que se encuentra entre la pared y la membrana celular).
El compuesto principal de la pared de las bacterias es el 'peptidoglicano', es un mucopéptido que aparece en casi todas las bacterias, y es único de este tipo de organismos, tiene además tres compuestos que no aparecen nunca en las células eucariontes. Estos compuestos son: T-aminoácidos, Acido murámico y Acido aminopimérico.
El peptidoglicano que confiere rigidez a la célula, si se elimina por una lisocima, en un medio hipotónico, determina la lisis del organismo. El peptidoglicano se compone de una cadena lineal de compuestos alternantes de N-Acetil-D-Glucosamnina y N-Acetil-Acido Murámico (NAM). Se forman cadenas separadas que se entrelazan en distintas zonas por puentes peptídicos cortos. Unidas al Acido Murámico existen unas cadenas laterales proteicas que contienen 4 aminoácidos en una secuencia específica, que es L-Alanil-D-Glulanil-X-D-Alanina. X, puede ser un aminoácido neutro como la homoseina, pero normalmente es un aminoácido básico como la L-Ornitina, L-Lisina ó el Acido Diaminopinético.
H NHCOCH3 CH2OH
¦_________¦ ¦_________O
/¦ ¦\ H H /¦ \
/ OH H \ ¦ ¦ / OH \ +-----+
O --+< >¦ ¦< >¦ ¦
¦ \ H / +--- O ---+ \ H / ¦ ¦
H \¦________O/ \_________¦/ H ¦
CH2OH /\ NHCOCH3 ¦
+------------------------------ O ------------+--+------------------+
¦ ¦ +-------------+ +
¦ +----------- O H ¦
¦ Enlace covalente ¦ ¦
¦ H NHCOCH3 CH2OH con cadenas adya CH CH3¦
¦ ¦_________¦ ¦_________O centes. ¦ ¦
¦ /¦ ¦\ H H /¦ \ H C - O ¦
¦ / OH H \ ¦ ¦ / OH \ ¦ +-----+ ¦ (1)
¦< >¦ ¦< >¦ ¦Ala-L¦ ¦
¦ \ H / +--- O ---+ \ H / +--- O ¦Glu-D¦ ¦
H \ ¦________O/ \_________¦/ ¦Lis-L¦ ¦
CH2OH /\ NHCOCH3 ¦Alg-D¦ ¦
(1) +-----+ +
En las bacterias Gram -, el peptidoglicano tiene probablemente una sola capa, y las cadenas no están muy enlazadas:
|
NAM - NAG - NAM - NAG \/\ NAM - NAG - NAM - NAG NAM - NAG - NAM - NAG \/\ NAM - NAG - NAM - NAG |
En las bacterias Gram +, la pared se compone de muchas cadenas mucopolipeptídicas, alrededor de unos 20 Bacillus Subtilus. Las uniones tienen lugar entre el grupo carboxilo del grupo terminal de D-Alanina y un aminoácido libre del tetrapéptido, o bien la unión se realiza entre dos aminoácidos terminales de una secuencia peptídica adicional. En las Gram +, las uniones entre las cadenas pueden situarse o por debajo o por enzima, dando mayor rigidez al conjunto, que entonces puede considerarse como una molécula.
Otro tipo que existe, son los 'Acidos Teicoicos', que son polímeros de azúcar fosfato, como el 'Glicerol-Fosfato' o el 'Ribitol-Fosfato', que se encuentran en las paredes celulares de las bacterias Gram +. Estos compuestos dan un medio favorable para la incorporación de iones, especialmente el Mg+2 ó también para la regulación de la actividad enzimática.
Los Acidos Teicoicos se pueden unir a enzimas autolíticas de la pared celular, y pueden participar en la regulación de las modificaciones líquidas que ocurren en el glucopéptido de la pared celular en la división.
Lipopolisacáridos :
Son parte fundamental de la pared de las baterias Gram -. Las bacterias necesitan este componente para ser patógenas, y en algunos puntos concretos de este compuesto, se realiza la absorción de bacteriófagos y bactericidas.
| Peptidoglicano + Ac. Teicoico. | |||
| Gram + | 40% a 50% | ||
| M. Externa = Lipopolisacáridos | |||
| Lipoproteinas | |||
| Gram - | + Lípidos | ||
| Peptidoglicano. 5% a 15% | |||
Los flagelos bacterianos poseen una disposición mas o menos helicoidal, y son varias veces mas largos que las células bacterianas. Se componen de subunidades de la proteina 'Flagelina', que aparecen al microscopio en cadenas de moléculas arrolladas en espiral. Los flagelos de distintas especies de bacterias se diferencian en la composición de aminoácidos, y en condiciones de acidez, los filamentos flagelares se pueden disociar en subunidades polipeptídicas. Por ejemplo, en Bacillus Punillus, poseen un peso molecular que oscila entre 30.000 y 40.000.
Si se eleva lentamente el pH, estas subunidades se vuelven a unir originando filamentos rectos. La forma recta de los filamentos se reorganiza para dar lugar a la forma ondulada más estable.
PH ácido -------------------------------> PH básico.
+-------------------------------------------------------------------+
Subunidades --------> Filamentos ----------> Filamentos helicoidales.
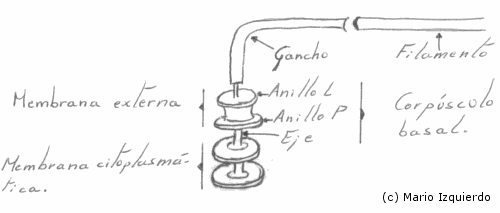
La estructura de cada flagelo se une a la membrana celular por una formación denominada 'Gránulo' ó 'Disco Basal', que posee una morfología específica de cada especie. El filamento flagelar se une a un gancho, y mediante este a un eje y a un conjunto de anillos en la pared celular y membrana.
El gránulo basal interviene en el transporte de energía desde el citoplasma o la membrana, hasta el flagelo.
El funcionamiento y montaje del flagelo es bastante complejo, por ejemplo en Escherichia Coli ó en Salmonella Tliphimurium, intervienen en el proceso unos 20 genes, incluidos en el que corresponde a la flagelina, y determinan el movimiento y dirección el flagelo. En general, los flagelos bacterianos giran rígidamente alrededor de su eje, en otras bacterias existe una posibilidad de deslizamiento sobre superficies sólidas, es un movimiento en el que interviene la secreción de mucílago.
Son unas estructuras muy frecuentes en las bacterias Gram -, y tienen un diámetro que oscila entre 5 nm y 10 nm. También en las bacterias existe un tipo de pelo especial que participa en la conjugación, y se conoce con el nombre de 'Pelo Sexual'. Su tamaño es mayor que el de los pelos normales, con un diámetro de 25 nm a 30 nm, y una longitud es intermedia entre la de los flagelos y la de los pelos normales.
En cada individuo sólo existen uno o dos pelos sexuales. Los pelos están compuestos de una proteina que se puede disgregar y reorganizar espontáneamente, siendo estos pelos huecos.
Durante la conjugación de las bacterias, el material genético pasa de la bacteria donante a la receptora, a través del pelo sexual. Los pelos son una zona de absorción de un grupo muy especializado de bacteriófagos, algunos de los cuales se absorben en el extremo y en otras zonas del pelo. Estos pelos intervienen en la adhesión de las células entre si, o las células con otras partículas del medio.
Tiene una estructura típica de unidad de membrana, pero el componente lipídico se diferencia del que hay en los eucariontes, porque contiene ácidos grasos de cadena ramificada que llevan ciclopropano, y raramente en esta membrana se encuentran ni esteroides, ni lecitina.
- Mesosomas :
Los mesosomas son invaginaciones de la membrana celular, que suelen estar asociados con el ADN de la bacteria, o con puntos de formación de tabiques en condiciones hipotónicas. Los mesosomas se evaginan y el material nuclear se arrastra hasta la membrana plasmática.
La estructura de los mesosomas es parecida a la de la membrana plasmática contigua, pero se diferencia de ella en la composición de los lípidos y proteinas incluidos en las enzimas. En determinadas bacterias, los mesosomas dan una superficie activa, mucho mayor que la que correspondería a la membrana plasmática, y es importante en los organismos que utilizan sustancias gaseosas poco solubles en H2O.
los mesosomas también facilitan el movimiento y participan en el mantenimiento de condiciones reductoras o anaerobias.
Estructuras similares a la de los mesosomas existen en bacterias fotosintéticas, y poseen forma de saco, tubulares o laminares, conteniendo todo el aparato fotosintético, incluidas las clorofilas, carotenoides y elementos de transporte de electrones, estas estructuras se conocen con el nombre de 'Tilacoides'.
Otras funciones de la membrana bacteriana, son:
· El mantenimiento de los gradientesosmóticos.
· El transporte de sustancias disueltas incluidas en el transporte activo.
· La organización de la síntesis de la pared celular y enzimas, que intervinen en la síntesis de sustancias precursoras de los polisacáridos de la pared.
· Participa en el metabolismo oxidativo y en la producción de energía.
Los procesos bioquímicos relacionados con la síntesis de ATP en las bacterias no fotosintéticas, es decir, la fosforilación oxidativa y la cadena respiratoria y también el transporte de sustancias e iones, se organizan en complejos multienzima proteina-cofactor. Algunas partículas que corresponden a la actividad de la ATPasa, relacionada con la fosforilación oxidativa, están distribuidas sobre la superficie interna de la membrana bacteriana.
En las bacterias no hay membrana nuclear, en Scherichia Coli, se ha demostrado por lisis de la membrana plasmática y marcaje con Timidina H Tritiada, que el ADN, es una molécula circular que puede presentar ramificaciones y posee una longitud de 1100 μm. Mientras que la célula de esta misma especie posee una longitud de 1 μm a 2 μm, otros organismos bacterianos se ajustan a este modelo general, no existe ninguna de las proteinas básicas, las histonas, que se asocian al ADN en las células eucariontes. En algunas bacterias hay ADN extracromosómico, que está en relación con elementos genéticos citoplasmáticos, entre ellos, los factores sexuales y los transferibles de resistencia a las drogas. Este ADN cromosómico normalmente está formando moléculas circulares, que se encuentran en el cromosoma bacteriano.
En algunos procariontes hay vacuolas llenas de gas que permiten al organismo regular su flotabilidad. En las bacterias fotosintéticas, en las rojas, en la cianofíceas y más raramente en las bacterias acuáticas no fotosintéticas, existe este tipo de vacuolas que pueden llegar a ocupar el 40% del volumen celular. Se encuentran rodeadas de una membrana proteica, de estructura distinta a la membrana plasmática. Además de las vacuolas existen en las bacterias inclusiones o gránulos con sustancias de reserva, e incluso en bacterias especializadas existe azufre molecular que se almacena en forma de cristales intracelulares.
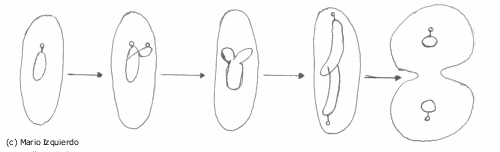
Los principales cambios citológicos que se observan son:
· El crecimiento celular.
· La división por formación de tabiques.
· Segregación en las células hijas.
· Escisión binaria o bipartición
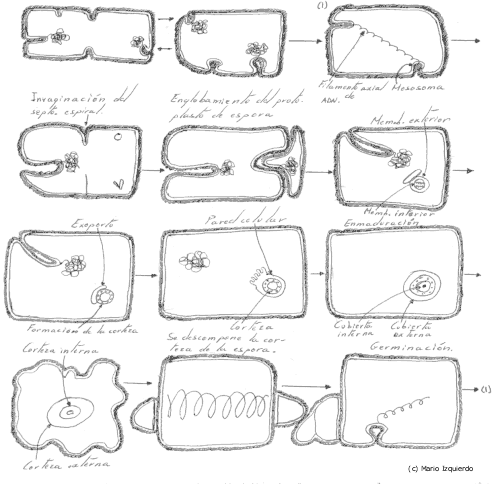
En la esporulación, la espora que se forma es muy diferente a la de la célula vegetativa, tanto por la forma como por la estructura, y además la espora es muy resistente a la radición y al calor. El proceso comienza con la formación de un filamento de cromatina axial, que es seguido de un proceso de invaginación de la membrana, formándose entonces una preespora que es englobada en la célula madre, y aparece rodeada de dos membranas. Entre estas dos membranas se sitúa y se sintetiza un material o corteza constituido por un mucopéptido. Cuando esta coteza está casi completa, una proteina que contiene una gran proporción de lisina y de cisteina, se condensa alrededor de la superficie exterior de la espora y forma la cubierta. Después, la espora madura forma una estructura muy refractaria que presenta a veces rugosidades y que se desprende por lisis de la corteza madre.
Esporulación y germinación del género Bacillus:
Las bacterias se pueden dividir de forma general por su aspecto morfológico. Las que poseen forma de bastón reciben el nombre de 'Bacilos', las esféricas 'Cocos' y las helicoidales 'Espiriles'. Las bacterias y cianofíceas se pueden dividir en dos grupos:
Estos dos grupos se colorean de forma diferencial por el método de Gram, descrito por Christian Gram en 1884. Es un método de coloración que utiliza dos colores: el violeta cristal y la safranina. Básicamente el violeta cristal tiñe las Gram +, y la safranina las Gram -. La división en Gram +, y Gram -, es una clasificación mas o menos natural, ya que las paredes de las Gram + poseen una composición diferente a la de las Gram -, aunque no es una clasificación natural desde el punto de vista evolutivo. Otra clasificación que se puede hacer, es la basada en los tipos existentes de dexosirribonucleótidos y sus proporciones relativas.
- Bacterias fotosintéticas:
Utilizan la energía solar para reucir el CO2, pero a diferencia de las plantas verdes no tienen como fuente de electrones el H2O.
Las bacterias sulfurosas, que pueden se purpúreas o verdes, utilizan el sulfuro de hidrógeno como fuente de electrones para sintetizar NADH y también ATP. Durante el proceso se forma azufre elemental, que puede almacenar como gránulos en el organismo.
Las bacterias fotosintéticas poseen clorofilas especiales asociadas a las membranas de los mesosomas. La mayoria son anaerobias, no toleran el oxígeno libre y se encuentran en el fondo de las zonas acuáticas, donde llega energía que no ha sido aprovechada en los estratos superiores.
- Bacterias quimioautotrófas:
Pueden formar hidratos de carbono a partir de materias inorgánicas, sin la intervención de energía luminosa. Las bacterias sulfurosas quimioautotróficas, utilizan sulfuro de hidrógeno para obtener energía y carbohidratos.
Las bacterias férricas oxidan los compuestos de hierro.
Las nitrificantes oxidan NH3 para obtener nitratos, y son muy importantes en el ciclo del nitrógeno.
Bacilos :
Entre los bacilos Gram +, se encuentra el género 'Clostridium', cuyos ejemplares son anaerobios. Algunas especies de este género producen enfermedades como:
· Clotridium Tetanium.- Produce el tétanos. Los bacilos de esta especie liberan una toxina que actúa sobre la sinapsis en el sistema nervioso.
· Clostridium Botulium.- Produce el botulismo. La toxina se libera en los alimentos mal enlatados y actúa al ser ingeridos estos alimentos, sobre el sistema nervioso y las placas motoras terminales.
Otros bacilos Gram +, además de los Clostridium, son:
· Bacillus Antracis.- Produce el antrax, que puede darse en animales domésticos y en el hombre.
· Lactobacillus.- Es un bacilo importante desde el punto de vista económico, ya que se utiliza en la producción de yogourth, queso, etc.
Entre los bacilos Gram - se encuentra 'Escherichia Coli', que es común en el intestino humano, y sintetiza la vitamnina K y algunas del grupo B. Otras Gram - son:
· Salmonella Typhi.- Produce el tifus.
· Vibrio Cholerae.- Produce el cólera. Los dos bacilos se pueden encontrar en el intestino humano, a donde llegan con el alimento o el agua contaminados.
· Yersina Pestis.- Se transmite por la picadura de una pulga de rata infectada. La presencia de este bacilo en cantidades suficientes, produce transtornos en el sistema linfático, y además produce la peste bubónica, o también puede afectar al sistema respiratorio denominándose entonces neumonía o peste neumónica.
Cocos:
Los cocos esféricos se suelen presentar formando agregados de estructura variable.
· Entre los Gram + destacan los estreptococos, que forman cadenas, y son responsables de las enfermedades que afectan al sistema respiratorio, infecciones en el oido medio, escarlatina y tiña.
· Los neumococos, crecen en pares de individuos y de ahí el nombre de diplococos. Es responsable de la neumonía bacteriana.
· Estafilococos.- Crecen formando paquetes planos. Son frecuentes en la piel, por ejemplo el Staphylococcus Albus y el Aureus. Si alguna de estas especies penetran en heridas, quemaduras o en alimentos en malas condiciones, pueden producir transtornos.
Entre los Gram +, existen dos grupos:
| Meningitis | |||
| Neisseira | Gonorrhoeae | ||
La primera especie produce la meningitis menincócica y la segunda la gonorrea.
Entre las bacterias de forma helicoidal están los espirilos y las espiroquetas. Las dos formas se encuentran en el agua, y un tipo de espiroqueta es responsable de la sífilis.
Otro grupo de procariontes son los 'Actinomicetos'.
Actinomicetos:
Se denominan así porque en principio se incluían en el grupo de los hongos, debido a su aspecto de filamento delgado, compuestos por muchas células. Después se comprobó que poseían una estructura de procariontes.
Juegan un papel muy importante en la descomposición de restos orgánicos en el suelo y su uso para la producción de antibióticos como: estreptomicina, cloranfenicol, tetraciclinas y otros tipos. También, a partir de los actinomicetos, se obtiene la 'actinomicina', utilizada en muchos experimentos relacionados con los ácidos maleicos, porque actúan sobre la replicación y transcripción del ADN.
Micobacterias y Corinebacterias.- Son parecidas a los actinomicetos. Entre las micobacterias se encuentran las responsables de la tuberculosis y la lepra. Entre las corinebacterias, están las reponsables de la difteria.
Micoplasmas.- Son bacterias inmóviles, sin pared celular e incluyen a los organismos de menor tamaño, entre las bacterias hasta 0,1 μm, y en general son esféricos, pudiendo tener vida libre o ser parásitos en plantas y animales. Entre ellos se encuentran los que producen la neumonía del ganado y la neumonía atípica.
Rickettsias y Clamidias.- Tienen un tamaño similar al de los micoplasmas, diferenciándose de ellos, porque todos son parásitos intracelulares. Entre los rickettsias están los responsables de la fiebre tífica, producida por la transmisión de un piojo. Entre las clamidias se encuentran los que producen la psitacosis y el tracoma.
Cianofíceas o algas verde-azuladas:
Son procariontes fotosintéticos con clorofila A, como las plantas, y utilizan el H2O como fuente de electrones para reducir el CO2 a hidratos de carbono. Pueden ser unicelulares o desarrollarse en filamentos con las células interconectadas, así como moverse por deslizamiento. Algunas algas verde-azuladas poseen células incoloras o heterocistes, que pueden fijar el nitrógeno atmosférico. El color de estos organismos se debe a la presencia de dos pigmentos que se presentan en la clorofila: la 'ficocianina azul' y la 'ficocianina roja', de la combinación de estos elementos resultan ser de color púrpura, rojo o negro. Viven desde aguas termales hasta el límite superior de marea, en costas marinas; en este caso la pared celular y la capa de mucílago las protegen de la desecación. Existen fósiles de cianofíceas de hasta 2300 m.a. que han sido encontrados en Sudáfrica.
En la transcripción y traducción en las bacterias, las moléculas de ARN-m se unen a los ribosomas al mismo tiempo que son transcritas, formándose ribosomas o polirribosomas en los que la información genética es inmediatamente traducida para la síntesis. La ARN-polimerasa se desplaza a lo largo del ADN, permitiendo la síntesis progresiva de ARN-m.

© Mario Izquierdo, 2024 | Aviso legal | Política de Privacidad